Introduction
The Middle Triassic limestones of the Luoping area were identified as an important source of abundant and exceptionally fossils in 2007 by Q.Y.Z. during regular mapping in Luoping County, near the east border of Yunnan Province (Fig. 1A). Abundant fossils were identified in rock exposures over a wide area, and it became clear that the limestone hills were the source. Local farmers showed visiting geologists examples of exceptionally well-preserved fishes and marine reptiles.
In 2009, Q.Y.Z. with a team of colleagues and students decided to perform an experimental excavation in the Shangshikan Hill (coordinates, N 24o46’47.6”, E 104o19’40.7”), 2 km northeast of Daaozi village, removing many tonnes of rock to expose the complete succession through Member II of the Guanling Formation. The logged section was about 45 m thick, including 80 numbered beds of light-gray limestone and dark, calcareous muds, many of them characterized by their fossil content. Initial reports (Zhang et al. 2008; Hu et al. 2011) announced the new site, termed a fossil lagerstätte or a fossil biota, and argued their importance in terms of the evolution of life.
Since 2007, further excavations have continued at the Shangshikan site, and many new fossiliferous sites have been logged and collected. By 2009, the geologists realized the new fossil assemblages were on a par with the other well-known Triassic marine biotas of this region across the Yunnan-Guizhou provinces border, including Panxian, Xingyi, and Guanling (Hao et al. 2006; Motani et al. 2008; Wang et al. 2008; Jiang et al. 2009, 2010; Lu et al. 2018). Collectively, these sites are world famous for the exceptional data they provide on the recovery of life from the devastating end-Permian mass extinction, and especially on the evolution of key marine groups including limulids, malacostracans, fishes, and reptiles (Chen and Benton 2012; Benton et al. 2013).
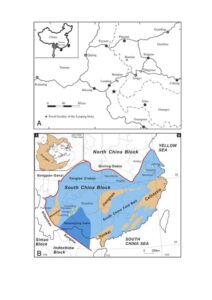
Figure 1. Topographic and geological maps. A) The location of Yunnan Province in China (top left) and the Luoping area, showing the site of the Luoping Geopark. B) Geology of South China in the Triassic, with general location of the South China Block in an Early–Late Triassic palaeogeographic map of the world (top left). Tectonic map illustrating major blocks of South China, interpreted suture zones and extent of the Nanpanjiang Basin and Yangtze Platform, with sites of named localities and biotas (yellow stars). ‘Old land’ is brown, shallow seas pale blue, and deep marine basins dark blue. B is based on a map by Jinyuan Huang in Benton et al. (2013).
Geological Background
Luoping is located towards the western end of the South China Block (Fig. 1B), which documents the north shore of the Tethys Ocean through more than 70 million years of Earth history from the Middle Permian to the Late Triassic. The block measures 3000 km wide and 1500 km from north to south. The deepest marine waters are documented in the Nanpanjiang Basin in Guizhou Province, and estimated water depths become shallower to the east, north and west. Around the Guizhou-Yunnan border, where the famous marine biotas of fishes and reptiles occur, water depths are shallow, based on the evidence of sedimentology and fossils, especially the occurrence of microbially induced sedimentary structures, trace fossils, abundant benthic organisms, and occasional wash-ins of terrestrial organisms. Further west in Yunnan Province, the fringes of the South China Block are reached where the Permian-Triassic sediments transition into fully terrestrial red beds.
The Triassic succession in the South China Block (Fig. 2) is especially important internationally because it documents such a long span of time with relatively complete and detailed documentation through long measured sections, some of them hundreds of meters thick (Benton et al. 2013). The maximum thickness is some 4 km from the Late Permian through middle–late Triassic (Wang 1985). This extraordinary volume of sediment, in terms of total area of outcrop and total thickness, means that there are great opportunities for detailed study.
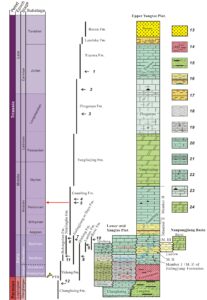
Figure 2. Schematic stratigraphic column illustrating the Permian and Triassic formations of the Yangtze Platform, with the age of the Luoping Biota marked by a red arrow. The different formations and biotas are dated according to biostratigraphy and radiometric dates, all compared with the international geological column. Numbered vertebrate-bearing units, from 1 to 12, are 1, Guanling biota; 2, Xingyi biota; 3, reptile layers below the Xingyi biota; 4, Panxian biota; 5, Luoping biota; 6, fishes and reptiles from the Nanlinghu Formation in Chaohu; 7, fishes from the Daye Group in Huangshi, and reptiles from the Jialingjiang Formation in Nanzhang and Yuanan, Hubei Province; 8, fishes from the Qinglong Formation in Jiangsu; 9, fishes from the upper Luolou Group in Guangxi Province; 10, reptiles from the Beisi Formation in Guangxi; 11, fishes from Helongshan Formation in Chaohu; 12, fishes from the upper Changhsing Formation in Meishan. Major lithologies, numbered 13–24, are 13, siliciclastics; 14, siliciclastics with mudstone; 15, siliciclastics with mudstone and coal; 16, mudstone, silt and fine sandstone with colourful mudstone and silt sandstone interbeded; 17, mudstone and silt sandstone with shale; 18, mudstone; 19, mudstone with limestone intercalated; 20, limestone with mudstone interbedded; 21, limestone and marl with mudstone interbedded; 22, worm limestone and limestone with bioclastics; 23, medium- and thick-bedded limestones with chert nodules and laminated or bedded; and 24, dolomite with dolomitic limestone interbedded. Based on an image by Jinyuan Huang in Benton et al. (2013).
The Luoping biota is found in the Guanling Formation, a unit exposed extensively over eastern Yunnan and western Guizhou provinces. The Guanling Formation at Luoping (Fig. 3) consists of carbonates and clastic rocks and is subdivided into members I and II. Member I is 333 m thick and is mainly mudstone and argillaceous dolomite with a volcanic ash bed (green pisolite) at the base. Member II is a 580 m thick succession of nodular limestone, silty limestone, micritic limestone and bands of dolomite. The overlying Yangliujing Formation comprises 1204 m of dolostone and dolomitic limestone interbedded with limestone breccias that show pseudomorphs of gypsum- and evaporite-solution breccias.

Figure 3. Sedimentary log through the Guanling Formation (Members I and II) and the Yanliujing Formation at the Shangshikan locality, near Daaozi village, Luoping county, Yunnan province, showing lithologies, sedimentary structures and key fossiliferous beds. Modified from Zhang et al. (2009).
The Luoping biota occurs mainly in a 16-m section of Member II of the Guanling Formation, beds 33–61 at Shangshikan (Fig. 3), as established during excavation of the hill (Fig. 4, 5). These are delimited at the bottom and top by thick-bedded, massive limestones and silty limestones, with high levels of bioturbation. These fossiliferous horizons of the Luoping biota are mainly thinly laminated micritic limestones alternating with thin- to moderately thick-bedded silty limestones. Lower units (Beds 2–66) are dark gray, medium- to thick-bedded, laminated micritic limestones with siliceous concretions, interbedded with a few thin ash beds. A highly diverse fish assemblage has been recovered mainly from Beds 33 and 44, and most saurichthyid fish fossils are found in Bed 48. Beds 57 and 61 yield marine reptiles, associated with shrimps, crinoids, and plant fossils. These units are followed by a 1.4-m-thick succession of dark gray medium- to thick-bedded nodular limestone intercalated with cherty concretions (Beds 67–74). The overlying 2.9 m of dark gray to yellowish-gray laminated muddy limestones (Beds 75–110) contain rare siliceous concretions. Numerous marine reptiles along with rare shrimps occur in Beds 78–81 and 106 (Fig. 3).

Figure 4. Excavating the Shangshikan Hill site, near Daaozi village, in 2009. Workmen are lifting slabs full of fossil fishes for the museum. Photograph by Q.Y.Z.

Figure 5. View across Shangshikan Hill, near Daaozi village, the key initial locality that yielded examples of the Luoping Biota, showing the layer-by-layer excavation, exposing extensive fossil-rich horizons, including fish-rich horizons on the main platform. The Chinese red flag stands at the top of the hill, and the custodian’s house in the foreground. This location has now been substantially altered to improve access for visitors, as a key site within the Geopark.
The fossiliferous, dark-colored micritic limestones of Member II of the Guanling Formation, as seen around Daaozi (Figs. 3–5), have been mapped over an area of some 200 km2 in the north-eastern parts of Yunnan Province, and crossing into Guizhou Province, through the mapping program of the Chengdu Center of China Geological Survey. Outside the Luoping area, Member II of the Guanling Formation is represented mainly by bioturbated, pale-colored micritic limestones like those in the lower and upper parts of the section around Daaozi. This suggests that the Luoping biota was preserved in a semi-enclosed intraplatform basin that experienced some anoxia, whereas more typical, well-oxygenated sediments were being deposited at the same time in other locations over the wider area.
It is crucial to fix the geological age of biotas such as Luoping. The Luoping Biota was dated early by Zhang et al. (2009) who identified conodonts as part of the fossil assemblage, which belong to the Nicoraella kockeli Zone, characterizing the Pelsonian Substage of the Anisian Stage (Middle Triassic). More recent astrochronological dating through the South China successions (Li et al. 2018) dates the Pelsonian to 245–244 Ma, and the Nicoraella kockeli Zone, equivalent to the Paragondolella bulgarica Zone, as in the lower half of that time span, dating more exactly to 244.14–243.29 Ma, based on measurement of the Guandao section in Guizhou Province.
The Panxian Biota (Hao et al. 2006; Motani et al. 2008; Jiang et al. 2009), known since 2003, is also Pelsonian in age, but marginally younger than Luoping, and the fauna is different, so a case has been made to distinguish the two as distinct assemblages (Hu et al. 2011; Benton et al. 2013). The Panxian Biota also occurs in Member II of the Guanling Formation, but it is important to realize that this geological unit is distributed over hundreds of kilometers and is thick, and the key location of the Panxian biota, Yangjuan village in Xinmin district, is 300 km east of Luoping. The Panxian biota consists of brachiopods, bivalves, and ostracods, as well as diverse fishes and reptiles, but none of the other arthropod groups, including the abundant crustaceans, that have been found at Luoping. Conodonts give the age of the Panxian fishes and reptiles as corresponding to the Nicoraella kockeli Zone, as at Luoping, However, the Panxian fossils occur stratigraphically higher, by some 50 m according to field mapping (Benton et al. 2013, p. 207). The general Pelsonian age of the Panxian biota is confirmed by a U-Pb age of 244 Ma (Wang et al. 2014).
Scientific Importance
The scientific importance of the Luoping localities lies in the fossils, which are abundant, diverse, and in many cases exceptionally preserved, showing often unpreserved features such as soft tissues. The fauna (Fig. 6) includes arthropods (crustaceans, millipedes, limulids; Fig. 6K), of which the crustaceans are the most common and diverse, represented by decapods (Feldmann et al. 2012), isopods (Fig. 6H), cycloids (Fig. 6E, mysidiaceans, conchostracans and ostracods. Several specimens of fossil horseshoe crabs show preservation of soft-tissue anatomical details including the chelicerae, five pairs of walking appendages, opisthosomal appendages with book gills, muscles, and fine setae, all of which confirm remarkable conservatism in the anatomy of these famous ‘living fossils’ (Hu et al. 2017).
Molluscs of the Luoping biota include relatively common bivalves and gastropods, and much rarer ammonoids and belemnoids. Although only two specimens of belemnoids have been found, both show good preservation of beak parts and arm hooks (Fig. 6G). Specimens such as these are very rare, with examples known otherwise only from the Middle Triassic Monte San Giorgio, the Early Jurassic Posidonia Shale, and the Late Jurassic Nusplingen Limestone of Germany and Switzerland.
Less diverse and less abundant are the echinoderms and brachiopods. Only a few specimens of crinoids, starfishes and sea urchins (Fig. 6I) have been found, perhaps reflecting the generally dysoxic to anoxic seabed conditions. Only one lingulid brachiopod has been found.
Fishes are important elements of the Luoping biota. So far, over 30 taxa have been identified, belonging to 13 families of actinopterygians, the Palaeoniscidae, Rhadinichthidaea, Saurichthyidae, Perleididae, Colobodontidae, Louwoichthyidae, Peltopleuridae, Kyphosichthyidae, Lashanichthyidae, Venusichthyidae, Platysiagidae, Marcopoloichthidae and Habroichthyidae. The Middle Triassic was crucial in actinopterygian evolution with the emergence of the neopterygians, comprising holosteans and teleosts, and Luoping samples diverse lineages, including a new genus of perleidid fish, Luopingichthys (Sun et al. 2009), Louwoichthys, representative of a new family and order (Xu 2021), the semionotids Sangiorgioichthys (López-Arbarello et al. 2011) and Luoxingichthys (Wen et al. 2012), the largest holostean, the ionoscopiform Robustichthys (Xu 2019), the durophageous colobodontid Feroxichthys (Xu 2020), and platysiagids Platysiagum, Helmolepis and Caelatichthys (Wen et al. 2019). In addition, the basal neopterygian Gymnoichthys (Tintori et al. 2010) and four new species of the saurichthyids Saurichthys and Sinosaurichthys (Fig. 6B) (Wu et al. 2009, 2011; Zhang et al. 2010) have also been reported from Luoping. Most of these represent genera that are common to Middle Triassic ichthyofaunas elsewhere, such as Monte San Giorgio in western Tethys.

Figure 6. Typical fossils of the Luoping biota. A) Ichthyosaur (LPV 30986); B) carnivorous fish Saurichthys yunnanensis (LPV 20881); C) eugnathid fish (LPV 11846); D) tooth of archosaur (LPV 31411); note serrated margin; E) cycloid arthropod (LPI 32637); F) coprolite containing fish scales (LPI 61753); G) belemnoid with preserved arm hooks (bottom) and beak parts (top) (LPI 61543); H) Protamphisopus baii, an isopod arthropod (LPI 61713); I) sea urchin (LPI 61701); J) Sinosoma lupingense, a millipede (LPI 61593); K) Yunnanolimulus luopingensis, a xiphosurid arthropod (LPI 61299); L) conifer plant (LPI 60148). Scales bars, (A) 4 cm, (B) 5 cm, (G) 2 mm, (C, I–L) 1 cm, (D–F, H, J) 0.5 cm, respectively. All specimens are deposited at the Chengdu Center of China Geological Survey (LPI = Luoping invertebrates; LPV=Luoping vertebrates).

Figure 7. At some horizons at Luoping fishes occur in dense accumulations. Here, researchers are mapping and measuring the density of occurrence of the fossils. Photograph by Q.Y.Z.
Unusual new finds among the fishes were two new genera and species of coelacanths, Luopingcoelacanthus and Yunnancoelacanthus, one of the former with two embryos inside that suggest viviparity (Wen et al. 2013). These coelacanths, like the modern Latimeria, may have been adapted to living in dysoxic conditions, as indicated for the Luoping basin.
Many of the Luoping fishes are still undescribed taxa. At least three layers with mass occurrences of fishes are found among the fossil-rich horizons (Figs. 2, 7), and such mass accumulations occur at certain horizons, sometimes with dozens of specimens per square meter over whole bedding planes. These suggest that populations were killed off by mass mortality events either through large volcanic eruptions or upwelling of stagnant, poorly oxygenated bottom waters. Evidence for volcanic eruptions comes from several ash beds, while the latter is supported by the alternation of gray, thick-bedded bioclastic limestone and dark, thin-bedded micritic limestone. Frequent volcanic eruptions might have triggered anoxia and bloom of cyanobacteria in oceans.
Most large fishes from the Luoping biota were carnivorous, as indicated by their dentition consisting of large and striated conical teeth alternating with much smaller ones, and from coprolites (Fig. 6F), and the largest was a basal actinopterygian, Pteronisculus, up to 2.9 m long (Ren and Xu 2021). The colobodont Feroxichthys was a durophage, using its blunt-topped teeth for grinding shells of mulluscs or crustaceans (Xu 2020).

Figure 8. Holotype of Nothosaurus zhangi (LPV 20167), partial skeleton showing vertebrae and ribs (top) and the paired dentaries (middle right). Photograph by S.X.H.
The marine reptiles from Luoping have also attracted considerable attention, including ichthyosaurs, sauropterygians (nothosaurs, pachypleurosaurs), rare archosaurs (Fig. 6D) and tanystropheids (Benton et al. 2013). Particularly notable is the giant nothosaur Nothosaurus zhangi, first described from this site (Fig. 8; Liu et al. 2014). The assemblage of marine reptiles is similar to that of the Anisian–Ladinian-aged Monte San Giorgio locality in Switzerland. Some reptiles, for instance, ichthyosaurs (Fig. 6A) represent top predators and so suggest full rebuilding of the ecosystem. A specimen of the pachypleurosaur Diandongosaurus showed damage to its hindlimb, evidence that it had been attacked by another predator (Liu et al. 2021). A very important discovery was a specimen of the large, long-necked archosauromorph Dinocephalosaurus that was pregnant with a relatively well-developed embryo inside (Liu et al. 2017); this was the first evidence of live birth (viviparity) in Archosauromorpha, a major group of reptiles including crocodilians, dinosaurs, and birds, were thought to be egg-layers (oviparity).
Trace fossils are common in the thick-bedded limestone layers, especially Arenicolites, Rhizocorallium and Thalassinoides, indicative of deeper portions of the shallow marine continental shelf (Feng et al. 2017, 2018; Luo et al. 2018b; Luo et al. 2019). Tracks by crustaceans (e.g., Sinusichnus, Spongeliomorpha, Thalassinoides) coincide with the relative abundance of body fossils. However, many horizons at Luoping lack trace fossils, or yield only simple, fine, horizontal burrows, Planolites (Luo et al. 2019). In addition, coprolites are common, and some were produced by predatory fish, as shown by included fish scales (Luo et al. 2017; Luo et al. 2018a). Most unusual are large paired front paddle marks, Dikoposichnus, made by nothosaurs punting along slowly and perturbing bottom muds presumably to flush out crustaceans and other food species out of hiding (Zhang et al. 2014). These important tracks were first found at Luoping (Fig. 9) and have since been identified in other marine Triassic sections, and providing evidence for a novel mode of locomotion, occasionally seen in modern sea cows.

Figure 9. Pair of forelimb punt marks made by a large nothosaur, like Nothosaurus zhangi, showing appearance and size when slab is lifted A), and reconstruction of the animal in life B). Photograph by Q.Y.Z.; artwork by Brian Choo.
Terrestrial components of the fauna include teeth from terrestrial archosaurs (Fig. 6D), a millipede, and rare plants. Only two or three specimens of millipedes (Fig. 6J) have been found, belonging to the new genus and species Sinosoma lupingense, one of very few millipede fossils from the Triassic worldwide (Huang et al. 2018). Most of the fossil plants are conifers (Fig. 6L), and the relatively good condition of the complete branches and leaves suggests that they were transported only a very short distance, maybe 10 km, into the Luoping basin from nearby coastal areas.
Moreover, abundant filamentous microfossils of cyanobacteria are also commonly preserved in microbial mats of the lower fossil layer of the Luoping Biota (Luo et al. 2013), and the underlying stromatolites in the geopark and surrounding areas (Luo et al. 2014; Huang et al. 2022). These microbes may have contributed to the exceptional preservation of the Luoping Biota, exhibiting the same preservational mechanism as the Ediacara Biota (Gehling 1999).
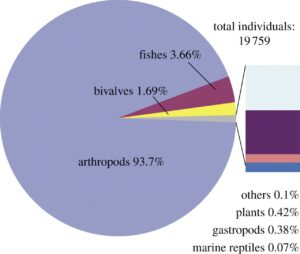
Figure 10. Pie chart showing the relative representation of the main macrofossils from Luoping, based on a count of nearly 20000 specimens. Rose, fishes; yellow, bivalves; purple, plants; blue, others; grey, arthropods; pink, marine reptiles; light blue, gastropods. Based on data from Hu et al. (2011).
In a summary of the 20,000 fossils initially collected from the Shangshikan locality, the Luoping biota (Fig. 10) is dominated by arthropods, which comprise over 90 percent of the fauna, followed by fishes (4%), then bivalves (2%), and then relatively small proportions (less than 1%) of plants, gastropods, marine reptiles and others. As summarized here, the fossils have been the subject of over 100 published papers so far, and these include important new discoveries that greatly enhance our knowledge of the arthropods, molluscs, fishes, and reptiles.
Geoparks in China
There are currently 284 national geoparks in China, and 41 of them have so far been accorded UNESCO international status, the highest number for any country in the world (Luan and Wang 2022).
Geoparks are just one way of ensuring conservation of geologically important sites. The main concerns of many conservationists are the plants, animals, and ecosystems of the modern world, and they might assume that the landscape, and especially the rocks, will preserve themselves. However, it was realized in many countries that the geological heritage will not survive unaided, as for example development and agriculture destroy unique geomorphological sites or quarries and coastlines are filled and covered in concrete, so obscuring important rock successions and fossil sites.
A geopark is defined as a delimited area that contains one or more geoheritage sites selected on the basis of scientific importance, rarity, scenic quality, or their relation to geological history, events, and processes (Eder 1999; UNESCO 2008). Wider goals are to conserve a healthy environment for the people living in and near the geopark, to educate the local people and people more widely in the earth sciences, and to foster sustainable, local economic development. Various international initiatives were drawn together in the 1990s, when the International Union of Geological Sciences (IUGS), with support from UNESCO, began to compile a global inventory of key geological localities and landscapes, leading to the establishment of the UNESCO Global Network of National Geoparks in 2001 (UNESCO 1999; Yang et al. 2011).
Much of the development of the worldwide geopark inventory happened at continental scale. The European Geopark Network was formed in 2000 (Zouros 2002; Yang et al. 2011). An Asia-Pacific Geoparks Network was promoted by Malaysia initially, and later by China and other nations (Yang et al. 2011) and the Asia-Pacific Geoparks Network is now well established, led by the Hong Kong Geopark in China. UNESCO Global Geoparks have been shown to have had positive economic impacts in Asia (Lee and Jayakumar 2021). Further, in the Americas, Argentina and Venezuela have advocated for the protection of geological heritage using geoparks. At the continental level, Europe and Asia rank first and second, with 81 and 65 UNESCO geoparks respectively, and then 13 and two UNESCO geoparks in the Americas and Africa. At the national level, China has the most UNESCO geoparks, with 41, and Spain ranks second, with 15. Six European countries (Italy, Britain, France, Greece, Portugal, and Germany) have a total of 38 geoparks. There are also 14 geoparks distributed in Japan and Indonesia (Luan and Wang 2022). These totals indicate the patchy development of the concept, with some nations forging ahead, and others following, and it may be some decades before there is even worldwide coverage, and all the key sites have adequate protection.
In China, interest in conserving geosites emerged in the 1980s, with a proposal in 1985 by Chinese geologists to establish geoparks to protect important geoheritage and permit scientific study (Xun and Ting 2003; Zhao and Zhao 2004; Luan et al. 2016). By 2004, China had 44 national geoparks, with one example being Yuntaishan in Henan Province, covering an area of 190 km2, and which was approved by the Chinese government in 2001. It received 600,000 visitors in 2001 which increased to 940,000 the following year, and this huge influx of visitors was accompanied by local employment and tourism infrastructure (Zhao and Zhao 2004). In 2004, China joined the Global Geopark Network (GGN) with eight of their 44 national geoparks accepted into this international framework. Also, in 2004, and as a measure of the official enthusiasm for this new concept, the Chinese Government offered to host the GGN central office in Beijing after the inaugural International Conference on Geoparks (Global Geopark Network 2018; McKeever and Zouros 2005). By 2020, China had 284 National Geoparks and 39 UNESCO Global Geoparks, rising to 41 in 2022.
The current total of 284 geosites or geoparks across the nation reflects these efforts since 1985, but with varying energy in different provinces, especially after the year 2000 (Yang et al. 2011; Luan and Wang 2022). The Chinese national geoparks include examples that are justified scientifically for their illustration of stratigraphy, structures, paleontology, mineral and ore deposits, geomorphology, hydrology, and environmental disasters. The sites cluster in central and eastern parts of China, with only a few sites in the far west of Tibet and Xinjiang and along the northern margins of the country (Luan et al. 2016; Luan and Wang 2022). The distribution of the geoparks reflects the underlying geology and terrain, but also human aspects such as the locations of previously funded geological heritage surveys and proximity to medium to large cities. There are lower correlations with economic development, capital investment, and research attention.
China was enthusiastic about the geoparks idea sometimes as much for developmental-economic reasons as for purely scientific, geological reasons (Ng 2014, 2017; Briggs et al. 2022). Geoparks were often located in remote areas and so could help improve local economies and alleviate rural poverty. The planning and execution could involve local communities, for example with the Hong Kong UNESCO Global Geopark, where Ng (2014) described how the government worked with local people and non-profit organizations to organize school visits, talks and seminars, and funded research to promote public awareness in geo-conservation based on scientific understanding.
Briggs et al. (2022) characterize the Chinese approach as “government-oriented.” They argue that the system has evolved through the adoption of sites already identified for their scientific or tourism potential by other government agencies. For example, the work of the regional branches of the China Geological Survey has identified many localities, including the current example of the Luoping Biota, as having geological significance. Alternatively, as described by Ren et al. (2013), tourism development potential might be recognized by the China National Tourism Administration (National 5-A rated tourism attractions), Ministry of Housing and Urban-Rural Development of the People's Republic of China (National Scenic Areas and UNESCO World Heritage Areas), and the Ministry of Land and Resources of the People's Republic of China (National Geopark and UNESCO Global Geoparks). After a possible site has been identified as a candidate for development, administrative committees are formed to coordinate across overlapping demands and interests, such as trying to balance geological research initiatives versus touristic development. Ren et al. (2013) note that the tourism aspects of newly established national geoparks generally develop very successfully, but the development of geoscience education programs is patchier in approach, and sometimes not at the standard required for UNESCO Global Geopark status.
In a special study of geoparks in Yunnan Province, Qi et al. (2017) distinguish sites like the Yunnan Stone Forest where visitors can immediately grasp the key points, from sites such as the Chengjiang Cambrian Biota geopark where the landscape says nothing and expert educational resources are required. The authors looked at display boards and other information available to non-expert visitors, and they criticize some of the explanations because they are full of unexplained jargon, and they particularly request that more attention should be given in the future to engaging professional science communicators to find the correct language and methods to transmit the correct scientific information in appealing and attractive ways to the public.
Steps to Establish Geopark Status for the Luoping Biota
Soon after the initial discovery of the Luoping Biota (Zhang et al. 2008, 2009, 2021; Hu et al. 2011), members of the China Geological Survey regional office at Chengdu who had conducted the mapping survey work, approached the Regional Development Office in Luoping city to suggest that the site was worthy of promotion as a geopark.
In 2014, Quarry 3, the biggest quarry of the Luing Biota National Geopark, was opened by the research team from Chengdu Center of China Geological Survey. A 60-m-thick section was revealed, showing the complete sequence of the interval yielding exceptionally preserved fossils. Some big slabs with large complex trace fossils Thalassinoides were recovered and kept in situ for visitors. In 2015, an expressway connecting the county town and the park was completed by the local government. A glass shelter covering the nothosaur tracks in quarry 2 was built to protect these unusual footprints (Fig. 11). In December 2016, a big sculpture with the handwritten message from the former prime minister Wen Jiabao (Fig. 12, 13), who is also a geologist, was constructed in the central area of the Luoping biota geopark.

Figure 11. Viewing area for the Dikoposichnus nothosaur tracks. The whole site was covered with thick glass sheets, and a safety fence installed, so visitors can examine the fossils close up. Photographs by Q.Y.Z.

Figure 12. Hand-written message from Wen Jiabao, the sixth Premier of the State Council of China, head of government from 2003–2013. The message reads ‘Luopoing Biota National Geopark; Wen Jiabao’(from the right to the left)


Figure 13. Marker block in the middle of the Luoping Biota National Geopark with Wen Jiabao’s calligraphy on the left. Photograph by Q.Y.Z.
The Luoping Biota site was approved as a national geopark in China on 13th November 2011, as one of 17 new geoparks accepted that day, the sixth batch to be approved nationally. The geopark has an area of 79 km2, with most of the geological heritage in the central zone occupying an area of 17 km2, and the
The geopark is divided into two major parts: the Daaozi area of the Luoping fossils and the Golden Chicken area of Karst Peaks, the former of which is characterized by the exceptionally preserved Middle Triassic fossils and the latter by spectacular karst landscape. Several additional places, such as the Jiulong Waterfall and Duoyi River, are also included in the national geopark.
In their evaluation of the geoheritage aspects of the site, Xie et al. (2013) summarize the scientific aspects as well as the tourism-related landscape features, including the Jiulong (‘nine dragons’) waterfalls nearby, the conical mountains of Triassic limestone standing steeply up from farmed fields, and the springtime yellow flowering of the canola crop (Fig. 5). They describe in particular the world-famous ‘cone peak landscape’ (Fig. 5) where as many as 5–10 white-to-gray-colored limestone peaks occur per square kilometer, each 50–200 m high, and emerging at steep slopes from the farmed fields full of brown soil and crops of canola and tobacco. Other geomorphological features are protected by the Geopark as well, including karst canyons, karst depressions, travertine beaches, and karst caves, and hydrological features such as springs, waterfalls and lakes.
In addition, Xie et al. (2013) report the aesthetic aspects of the Geopark, including the highly attractive, exceptionally preserved fossils, especially the abundant fishes, the unique and beautiful karst landforms, and the wider beauties of the landscape including such established tourist attractions as the canola blossoms in February and March each year, coinciding with the annual Spring Festival. As Xie et al. (2013) note, in 2002, the golden canola field blooming was awarded the title of “largest natural garden in the world” by the Guinness headquarters of Shanghai Great World, and in 2005, the Jiulong Waterfall Group was rated as “the most beautiful waterfall in China” by the Chinese Geographical Society, and Luoping Peak Forest was rated as “the most beautiful peak forest in China with significant aesthetic value.”
Finally, Xie et al. (2013) note five aspects of touristic development in the Luoping Biota National Geopark:
(1) Luoping includes leading national industrial and agricultural tourism sites, and has received many awards, including designation as one of China's excellent tourism counties, one of China's cultural tourism counties, one of 23 key scenic spots in Yunnan Province, one of seven scenic spots with development potential and international competitiveness, a provincial-level key scenic spot, and a national AAAA-level scenic spot.
(2) Luoping is an important part on the tourist trail through Yunnan Province, being located in the golden tourist area of southeast Yunnan, and only 157 km from the Stone Forest Global Geopark and World Natural Heritage Site; it has convenient transportation and outstanding geographical advantages.
(3) Luoping tourism resources are rich in science, knowledge, interest and appearance. The types of resources are rich and diverse, and relatively concentrated, which can be combined to meet the needs of different tastes, levels, and hobby groups.
(4) Luoping has a rich cultural heritage. The development of tourism, led by such locations as the Jiulong Waterfall, Duoyi River, and the sea of canola flowers, has enhanced the cultural sense of identity of local people, their pride and love of their environment.
(5) Luoping's rich geological heritage resources, plus strong ethnic customs, long history and culture, and splendid human landscape, constitute beautiful, harmonious, unique and irreplaceable high-grade tourism resources, with great tourism development potential and value. Luoping tourism has become an important pillar in the local economic development, leading to changes in planning for local social and economic construction.
Further Development of Geoconservation Aspects
Ten years after the approval of Luoping as a National Geopark, the infrastructure construction is almost finished. With the new expressway, the park is now easily accessible from the county town, and the entrance is marked with a remarkable, large sculpture (Fig. 14). A large new museum showing typical fossils of the Luoping biota will be available at the end of this year. New education programs for the public are in preparation, to tell the scientific story of the Triassic fossils in attractive ways. Meanwhile, more papers about the Luoping fossils have been published in recent years, especially on the fishes, making the Luoping Biota National Geopark one of the most diverse fossil sites of both vertebrates and invertebrates in the world.

Figure 14. Bronze marker at the entrance to the Luoping Biota National Geopark on the new expressway. The sculpture shows a traditional Chinese vessel, covered with a sculpted scene of the seabed with neopterygian fishes, and nothosaurs forming handles on either side. Photograph by S.X.H.
The next step for the Luoping Biota National Geopark is to seek international status as a Global Geopark and World Natural Heritage Site. To apply for admission as a World Natural Heritage Site, all the most important Triassic fossil sites from South China may be entered together. South China is not only famous for the GSSP (Global Stratotype Section and Point) type locality of the Permian-Triassic boundary at Meishan but is also well known for the exceptionally preserved fossil Lagerstätten from Early to Late Triassic, making this area one of the best places to study the recovery and radiation of marine life after the end-Permian mass extinction. This area preserves a complete stratigraphic succession of the Permian-Triassic transition, which developed in various facies, ranging from shallow to deep marine.
So far, more than ten fossil assemblages with exceptional preservation have been recovered from the Triassic of South China, including the Early Triassic Chaohu fauna (Yao et al. 2021) and the Nanzhang-Yuan’an fauna, the Middle Triassic Luoping biota and the Xingyi biota, and the Late Triassic Guanling biota. Highlights of recent discovery include the earliest ichthyosauriform from Chaohu, the filter-feeding marine reptiles from Luoping, and the first flying fish from the Xingyi. These kinds of discoveries were unexpected and have changed our perspective on the importance of the marine fossiliferous units in South China in documenting a complete remodeling of the marine ecosystem providing the roots of the modern fauna, an early start of the Mesozoic Marine Revolution, and evidence for a complete re-ordering and substantial energy and complexity increase in marine systems (Benton and Wu 2022). These are good reasons that all these sites of exceptional preservation of Triassic marine fossils together would justify future status as a World Natural Heritage Site.
Problems:
The vivid, exceptionally preserved fossils of the Luoping biota attracted many fossil collectors worldwide, and led to private collecting and smuggling in recent years. According to the Chinese law, all fossils with significant scientific value are under protected and private collecting is forbidden. Fossil sites within the national geopark are well protected by local government, but few localities outside the national geopark are excavated by private collectors occasionally. Moreover, the fossil sites of the Luoping biota are scattered in a large area more than 200 km2. Some localities are distributed in the neighboring counties, far from the boundary of the national geopark, making the fossil protection a tough task. Unlike the famous Triassic fossil sites from west Tethyan (e.g. Monte San Geogio, a world heritage site from the boundary area between Italy and Switzerland), which are located in a remote area and difficult to access, the Triassic fossil sites in South China are widely distributed, and are easily accessible. This is an advantage for scientific research but a disadvantage for fossil protection. In order to protect the Luoping fossils in a better way, more works are definitely needed.
A total of 230,000 visitors have been recorded since the approval of the Luoping biota National geopark. However, the number of visitors decreased dramatically in the recent three years due to the widespread coronavirus. It will rebounded quickly if the epidemic is finished. The official opening of the new facilities, including a new, modern museum, a tourist center, a television hall, and the parking area, will also attract more visitors. However, some the facilities for tourists are still not accomplished as planning due to the influence of the coronavirus. As mentioned above, some the of explanations on the display boards are full of unexplained jargon, and it is necessary to find the correct language and methods to transmit the correct scientific information to the public in attractive ways.
Acknowledgments
We thank Zhong-Qian Chen and an anonymous reviewer for very helpful comments on the MS.
Conflict of Interest
The authors declare that they have no conflict of interest in this work.