Daedalus: A Trace Fossil Expression of Complex Behavior
Daedalus is among the most complex and least understood Lower Paleozoic trace fossils. Despite that, its architecture is well known from the work of Sarle (1906) and Lessertisseur (1971). Daedalus is large, vertical spreiten (backfilled) structure, composed of J-shaped burrows whose displacement twists spirally inwards. Spreiten surfaces, resulting from the repeated translocation of the vertically oriented burrow hundreds of times, may cross themselves or other similar structures. They started formation at the water-substrate interface from an (almost) fixed central point and may pass down through one or more horizons and reaching as much as ~1 m deep, resulting in a succession of helicospiral, conical or spindle-shaped bodies.
Daedalus is a characteristic Lower Paleozoic trace fossil, particularly common in the Ordovician, high-paleolatitude shallow water sandstones of the “Armorican Quartzite” Formation and similar facies in France, the Iberian Peninsula, North Africa, the Middle East and Argentina (Rouault 1850; Delgado 1885; Lessertisseur 1971; Beuf et al. 1971; Seilacher 1964, 2000, 2007; Durand 1985; Poiré et al. 2003; Gibert et al. 2011; Heward et al. 2019). Daedalus not uncommonly occurs in high densities often completely dominating the physical structure of beds (Neto de Carvalho et al. 2016). The different ichnospecies of Daedalus, each developing characteristic, almost monoichnospecific ichnofabrics in the Lower-to-Middle Ordovician “Armorican Quartzite” of the Iberian Peninsula, are differentiated by the size and shape of the generating tube, and how these tubes are bundled into a single complex burrow: the type ichnospecies D. desglandi (Rouault) possesses complex helicoidal spindle patterns produced by descending translocations of a J-shaped tube, whose distal end was turning back towards the center (Lessertisseur 1971; Seilacher 2000). In D. desglandi the tube is relatively wide (10-20 mm) and forms bodies with an irregular, apparently chaotic distribution of burrow translations, with numerous intersections of previously exploited areas that usually expand in curvilinear bands defining the outer expression of the burrow complex until moving downwards and starting once again from the central axis. In Daedalus halli, the tube is narrow and straight (usually less than 5 mm wide, up to 10 mm), showing no evidence of active filling, and its displacement forms smooth carpet-like walls. A cone-like, helical 3D shape may be defined by a change of level in the sediment and the comma-to-spiral/circular shape of the spreiten on the bedding plane.
The producers of Daedalus are unknown and unlikely ever to be found, but were probably “worm-shaped” animals that opportunistically exploited the substrate for food. The distribution pattern and morphology of D. desglandi suggests they harvested sand-enveloping biofilms as a possible feeding strategy (as suggested for D. halli by Noffke, 2012). A long, worm-like animal burrowing downwards must have been capable of sediment displacement from one flank of the body to the other to produce a lateral backfill (Seilacher 2000). A possible process to explain the occurrence of Daedalus in clean quartz sands is the “subtidal pump” model of Riedl et al. (1972). In energetic shorelines or shallow marine bottoms frequently disturbed by storms, breaking waves act as a pump and bring down oxygen and nutrients in deeper levels of the substrate (Bock and Miller 1995). They also reduce the diversity of benthic life in shallower tiers in high-energy environments. The clean sands that constitute the quartzites may have acted like a sieve, and only fine organic particles would have reached the deeper tiers. This, together with the meiofauna that exploited the same food source among the sand grains, and the grain enveloping biofilms might explain why such complex burrows were built (Neto de Carvalho et al. 2016).
In the Iberian Peninsula, the Villuercas-Ibores-Jara and Naturtejo UNESCO Global Geoparks, located in the southern part of the Central Iberian Zone, in eastern Extremadura, Spain, and central Portugal, respectively, are territories with outstanding paleontological heritage. Among these, Daedalus mega-ichnosites are found in the “Armorican Quartzite”, corresponding to quartzite beds with dense D. desglandi ichnofabrics showing exceptional preservational and paleoecological features that can be followed for hundreds of meters or sometimes kilometers, with great lateral homogeneity. For these reasons, they were included as geosites in the geological heritage lists of both geoparks (Barrera & Gil Montes 2013; Cortijo et al. 2016; Neto de Carvalho et al. 2016; López Caballero et al. 2018; Fig. 1).
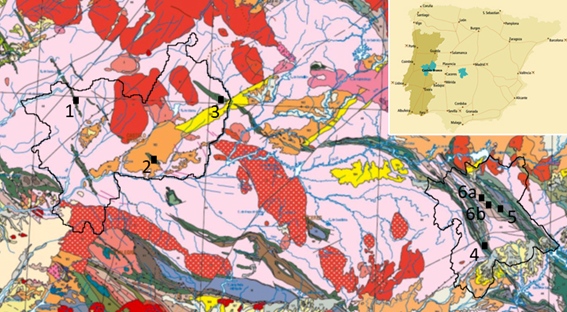
Figure 1. Location of D. desglandi geosites identified in Naturtejo and Villuercas-Ibores-Jara UNESCO Global Geoparks in the Lower-to-Middle Ordovician “Armorican Quartzite”. 1) Muradal-Fajão syncline. 2) Monforte da Beira. 3) Penha Garcia syncline. 4) Las Amoladeras. 5) Risco Carbonero. 6a and 6b) Camorros de Castañar de Ibor and Navalvillar de Ibor. Limits of both geoparks are placed on extract from the Geological Map of the Iberian Peninsula, Balearic and Canary Islands 1:1.000.000, 2015 edition of IGME/LNEG (all rights reserved). The symbols for the main stratigraphic units can be consulted at:
http://info.igme.es/cartografiadigital/datos/geologicos1M/Geologico1000_(2015)/pdfs/EditadoG1000_(2015).pdf
Daedalus Ichnofabric: Evolutionary Paleoecological Implications and Paleogeographic Distribution
An ichnofabric includes all structure and textural changes of the sediment resulting from bioturbation (and bioerosion) at all scales (Reineck 1963). The abundance and distribution of burrows reflect the non-linear sorting effects of physical and biological parameters, resulting in a disturbance regime at different degrees of patchiness in the colonization of the substrate. This is particularly true for the colonization of sessile to semi-sessile filter-feeding benthic fauna, or by vagile organisms with complex feeding strategies and/or cosmopolitan habits. Dense Skolithos ichnofabrics, known as piperocks, are usually considered to record the advent of deep burrowing by coelomate metazoans (possibly polychaetes or phoronids) during the Cambrian radiation (Droser 1991; Mángano & Buatois 2011). They first appear in the Early Cambrian in high-energy, nearshore, and storm-related siliciclastic deposits. The decline of the Skolithos piperock corresponds with the Ordovician faunal diversification and tiering complexification (Thayer 1983; Desjardins et al. 2010; Mángano & Buatois 2011, 2015; Liang et al. 2012). Together with Skolithos, which is also a common ichnofabric in the “Armorican Quartzite”, Daedalus may be seen as a pioneering product of the Cambrian substrate revolution (Early Cambrian forms described by Desai et al. 2010), a complex burrow that deeply penetrates the substrate for food processing and directly contributes to changing physical and chemical gradients in the sediments, similar to Dictyodora Weiss and Syringomorpha Nathorst. However, the general feeding behavior represented by Daedalus may have reached its evolutionary climax during the Early Ordovician. Dense Daedalus ichnofabrics are highly pervasive in the “Armorican Quartzite” and similar peri-Gondwanan, high latitude siliciclastics from Portugal (Delgado 1885; Neto de Carvalho et al. 2016), Spain (Gutiérrez-Marco et al. 2017) and Oman (Heward et al. 2019). These ichnofabrics occur in facies corresponding to very shallow marine, drifting sand bodies to lower shoreface and high hydrodynamic conditions (Figs. 2A, 3A), like amalgamated storm beds with planar lamination or hummocky cross-stratification.
To analyze the patch dynamics of trace fossils as a result of environmental disturbance, multifractal spectrum analysis was used as a measure of spatial ichnofabric heterogeneities in dense D. halli ichnofabrics from the Armorican Quartzite (Neto de Carvalho 2003; Neto de Carvalho and & Baucon 2013). The magnitude of the Daedalus ichnofabric fluctuations for the two stratigraphic sequences (Martim Preto and Serapicos in the NE of Portugal), both in area and in recurrence along the succession, shows that sandflat substrate colonization by the D. halli producer after each storm event was opportunistic, mostly multigenerational, with an exclusive and significant occupation of emptied ecospace (Neto de Carvalho and Baucon 2013). The patchy distribution pattern suggests a similar model of ecospace colonization to explain also D. desglandi architecture based on the subtidal pump mechanism of interstitial water exchange by wave action.
At Naturtejo UNESCO Global Geopark, Neto de Carvalho et al. (2016) described mega-ichnosites composed of quartzite beds with dense D. desglandi ichnofabrics which, in certain circumstances, can be followed for kilometers (Fig. 2). The Penha Garcia Formation (seen at site 3 in Figure 1), a formal regional name for the “Armorican Quartzite” at the Penha Garcia Ichnological Park (see this volume), is classified into two main groups of facies associations, which are interpreted as deposited in a mixed asymmetrical delta with along-strike variations between wave-dominated strandplain (updrift) and river-dominated deltaic settings (downdrift) (Bayet-Goll & Neto de Carvalho 2020). In the wave-dominated facies, D. desglandi beds are occasionally found in tide-influenced distributary channel fills at the delta front (Bayet-Goll & Neto de Carvalho 2020). The preservation of only one behavioral strategy, the substrate depth affected by these structures, the high density of burrows, and passive patchiness rates are all characteristics of r-selected populations. Such large-scale and frequent disturbance events as flood discharges at the delta front or storms (with remobilization and redistribution of sediments, endobenthos, and particulate food) made these Daedalus ichnofabrics one of the earliest opportunistic behaviors in the fossil record. They are also found in similar facies at Villuercas-Ibores-Jara UNESCO Global Geopark, where Daedalus beds can be followed for hundreds of meters (Figs. 3A–C; see also Cortijo et al. 2016), showing one of the best preservations known for this ichnogenus, and even including behavioral deviations to the normal architecture (Figs. 3D-G).

Figure 2. Muradal-Fajão syncline where the main Daedalus mega-ichnosite in the Naturtejo UNESCO Global Geopark is found. A) Basal bed soles crowded with D. desglandi at Cardal, Muradal Mountain. B) The same bed as A at Santa Luzia dam, 20 km NW from Cardal and showing the same dense ichnofabric. C) Cross section of the bed in B showing D. desglandi disrupting completely the primary fabric. Almost all the overlying beds show crowded D. desglandi ichnofabrics. D) Bed sole view of the D.desglandi ichnofabric at Portelo geosite, showing the dense distribution of spirally coiled burrow complexes, up to 30 cm wide (see Neto de Carvalho et al. 2016).

Figure 3. Risco Carbonero, the main Daedalus mega-ichnosite from Villuercas-Ibores-Jara UNESCO Global Geopark. A) General view of the succession exposed on the top of the mountain. B) Bedding plane showing the typical dense ichnofabric. Black scale is 15 cm. C) D. desglandi, with different spindle-shaped burrow complexesin cross section, evidencing the exceptional preservation of these trace fossils at the geopark. D) Cross section parallel to bedding of one of the D. desglandi burrow complexes showing the chaotic filling of the available ecospace with successive protrusive and outward expanding burrow bands. E) Dense vertical burrowing with many intersections of previously utilized areas. Coin as scale is 23 mm. F) Sediment disruption heterogeneities produced by burrow translocation and changing of tier, highlighted by post-lithification oxidation. G) Deviation of the helicospiral coiling behavior to a Zoophycos-like pattern.
Daedalus Mega-ichnosites: Geological Heritage of International Relevance
Daedalus desglandi records the feeding complexity of a highly successful pioneer animal in the early phase of ecological expansion into a non-uniformitarian marine environment (McIlroy & Garton 2010). Neto de Carvalho et al. (2016) reviewed the occurrence of the ichnogenus Daedalus in Portugal and compared it with other known sites all over the world. Although a relatively common trace fossil in high-latitude quartzite successions during the Early Ordovician to the Early Silurian, the representation of typical Daedalus ichnofabrics in large scale exposures was mostly unknown. A site in the “Armorican Quartzite” of the NE Portugal, locally known as the Pedras Escrevidas (“Written Stones”) shows intense bioturbation covering a 40 m2 surface mostly consisting of D. halli with the typical “Humillis”, spreiten spiralling preservation shown in bedding plane intersections. The site was described in Neto de Carvalho & Rodrigues (2002, 2003), Sá et al. (2002), Neto de Carvalho (2003), Gutérrez-Marco et al. (2007), and Neto de Carvalho & Baucon (2013). In the Lower Ordovician of the Oman mountains, a marker bed of regional stratigraphic significance in the Amdeh Formation is known for the dense ichnofabrics of D. desglandi and D. labechei (Seilacher 2007; Heward et al. 2019) and can be traced for kilometers. At Naturtejo UNESCO Global Geopark, Neto de Carvalho et al. (2016) described basal beds of the “Armorican Quartzite” which are Early-to-Middle Ordovician in age, with large exposures of the D. desglandi ichnofabric that can be followed, with interruptions, for at least 6.5 km. These D. desglandi beds are known now to reach at least 20 km in extent (Fig. 2), beyond the territory of the geopark. In the same formation, but at Villuercas-Ibores-Jara UNESCO Global Geopark, Barrera & Gil Montes (2013), Cortijo et al. (2016) and Neto de Carvalho et al. (2018) identified geosites with D. desglandi crowded beds, including the outstanding succession of Risco Carbonero, which shows some of the best-preserved forms of large Daedalus presently known, and wide bedding planes exhibiting their complex geometries. Even the smallest details of this ichnospecies architecture, including also behavioral deviations from the normal pattern, can be found here (Fig. 3). Finally, and not far to the east of the Villuercas-Ibores-Jara Geopark, Gutiérrez-Marco et al. (2017) found a large area at Fontanarejo de los Montes (Ciudad Real), where the “Armorican Quartzite” bedding planes show very crowded ichnofabrics, composed of well-preserved small and deep spindle-shaped D. desglandi, with up to five whorls.
The sedimentation rates and time available for colonization between sedimentary events determined overall density and spacing of the Daedalus ichnofabric. Moderate sedimentation rates, coupled with minor scour erosion, allowed multiple colonization events of multiple generations of D. desglandi producers to occupy the same substrate in the lower shoreface, sand sheet complexes, similar to the Skolithos piperock (Desjardins et al. 2010). This complex feeding behavior developed in deep tiers and adapted to soft-, bioturbated and shifting substrates, may not have been able to cope with the diversification of actively pumping filter-feeders and tiering complexification during the Ordovician, together with the progressive thickening of the mixed layer, and became extinct before the Middle Silurian. Daedalus behaviou had its biogeographical burst during the Early Ordovician (Seilacher 2000), in the “Armorican Quartzite” and similar facies but declined towards extinction during the GOBE (Neto de Carvalho et al. 2016). Thus, and in conclusion, the Daedalus beds from Naturtejo and Villuercas-Ibores-Jara UNESCO Global Geoparks are outstanding examples of paleontological heritage of international relevance for their kilometric extension, constituting mega-ichnosites, excellent preservation and for their evolutionary paleoecological implications which help us understand early metazoan radiation in marine substrates. Their importance and the ease of access to several of these mega-ichnosites meant they were listed as geosites in both UNESCO Global Geoparks and included in sustainable development projects.
A Cross-border Geotourism Cooperation Project Between Naturtejo and Villuercas-Ibores-Jara UNESCO Global Geoparks
Daedalus mega-ichnosites are important geosites of Naturtejo UNESCO Global Geopark in central Portugal and Villuercas-Ibores-Jara UNESCO Global Geopark, central Spain (Barrera & Neto de Carvalho 2018; Neto de Carvalho et al. 2018). In Villuercas-Ibores-Jara UNESCO Global Geopark, Daedalus can be examined in several interpretive centers and geosites (Figs. 3, 4A, C–D). Among these, the Risco Carbonero geosite offers the possibility to examine D. desglandi in loose blocks, bedding planes and sections in a setting with a spectacular view of an “Appalachian” landscape of tightly folded elevated synclines (Fig. 4B). The Cerro de las Amoladeras geosite offers particularly good lateral views of dense aggregates of D. desglandi. This trace fossil and the visit to its geosites are included in both educational programs and tourism offers (Cortijo et al. 2016).
In the Naturtejo UNESCO Global Geopark, beds extensively bioturbated with D. desglandi can be followed for several kilometers along the Muradal mountain through the Portuguese section of the well-known International Appalachian Trail (IAT). Officially called GR38 “Grande Rota do Muradal-Pangeia”, it is the first section of the International Appalachian Trail, recently opened in Portugal. Plans are underway to connect, by trails and tracks, the Muradal mountain, throughout the geopark, with the Spanish sector of IAT, which also crosses Villuercas-Ibores-Jara UNESCO Global Geopark. The GR38 offers views from the summit crests of the Appalachian-type landform where Daedalus beds stand out, including the interpretation of its and other geosites. One of the main starting points of the trail is the village of Estreito, where the local main church was built with quartzite rocks brought from Muradal mountain, and where the heraldic symbol of the town was long ago placed in a Daedalus rock (Fig. 4E). The trail became a popular adventure in central Portugal, despite the wildfires that caused great devastation in the summers of 2017 and 2020. Nevertheless, the diversity of themes along the trail, and the interpretation provided, mean this sector of the IAT has great potential as an educational tool (Figs. 4F–G). Daedalus was also featured in the film The Importance of Being Hard which was a production of the Spanish film company Libre Producciones, with the support of Villuercas-Ibores-Jara and Naturtejo UNESCO Global Geoparks. The premiere of this documentary about the evolutionary implications of the mineralized skeleton was in 2019. The documentary is centered on Cloudina, the earliest fossil with a mineralized shell and the main paleontological feature of Villuercas-Ibores-Jara UNESCO Global Geopark (see this volume). In Naturtejo UNESCO Global Geopark, Daedalus at the Portelo geosite was included in the movie as an example of early soft-bodied organism activity in the colonization of marine benthic ecospaces (Fig. 4H).

Figure 4. Daedalus as educational resources in Villuercas-Ibores-Jara and Naturtejo UNESCO Global Geoparks. A) Educational tools and fossils available at Visitors’ Reception Center in Cañamero village, Villuercas-Ibores-Jara Geopark. B) Interpretative panel at the Risco Carbonero geosite explaining the presence of Daedalus beds. C) The hall dedicated to the paleontological heritage of Villuercas-Ibores-Jara Geopark at the Visitors’ Reception Center in Cañamero. D) Interpretative panel about marine life during the Ordovician period depicting D. desglandi. E) The symbol of the town of Estreito in the public square, Naturtejo Geopark. F) One-day field visit for students of the 12th grade of a local high school hiking along the IAT at the Muradal mountain. G) Interpretation of Daedalus producer behavior and paleoenvironment in the artwork of Andrea Baucon “Tracemaker”. H) La Importancia de ser Duro, making of the documentary by the Libre Producciones team at the Portelo geosite, Naturtejo Geopark.
The “Armorican Quartzite” is a lithostratigraphic formation that is especially well represented in the landscape of the province of Cáceres (Spain) and in the Beira Baixa region (Portugal). It is visible at the top ridges of the mountain range that connects Penha Garcia to Guadalupe along a line that joins Naturtejo (Portugal) and Villuercas-Ibores-Jara (Spain) UNESCO Global Geoparks (Fig. 1). These geomorphological, Appalachian-type landforms constitute the Bridge over the Armorican Quartzite which is solidly supported in important natural areas of high scenic value in both geoparks, besides the Canchos de Ramiro and the sierras of Cañaveral Natura 2000 sites and the Monfragüe and Tajo-Tejo International Biosphere Reserves.
The project is the result of the continued collaboration of both geoparks and their communities that have considerable experience in cross-border cooperation projects. The geoparks cooperation is included in the framework of REDTI, CBC PROGRAMME SPAIN-PORTUGAL 2007-2013 (POCTEP), also known as Interreg Spain-Portugal. The Bridge over the Armorican Quartzite project is based on several pillars of activities:
Tourist pillar - Promotion of the Route (the “bridge”) linked to the Armorican Quartzite mountains between Villuercas-Ibores-Jara and Naturtejo UNESCO Global Geoparks (Figs. 5A, C). The Armorican Quartzite Route will take advantage of existing trails and other attractions (Fig. 5B) thanks to the activities of Naturtejo, the local LEADER groups and Villuercas-Ibores-Jara. Information and interpretative equipment will be created (Figs. 5C, D).
Promotional pillar - Promotion will be carried out through multiple channels, on the webpages of both geoparks as well as various social networks. A promotional book and a tourist guidebook in Spanish, Portuguese and English will be printed. Video in a variety of formats for use in exhibition rooms, websites, events, etc. will be produced for display in specialized local and international trade show fairs.
Educational pillar - Both geoparks work tirelessly to integrate the knowledge of the territory in classrooms of local schools but also nationwide. The project will serve to share methodologies, design common didactic units about the Armorican Quartzite and carry out practical activities in formal and informal education.
Scientific pillar - The scientific teams of both geoparks will establish guidelines for the correct interpretation of the Armorican Quartzite Route and will design lines of common work, for example, those referring to Paleozoic paleontology.
International pillar - At a global level, the project will be disseminated in the UNESCO Global Geoparks through the Global Geoparks Association. The connection with the International Appalachian Trail association will be an important asset.
Naturtejo and Villuercas-Ibores-Jara UNESCO Global Geoparks offer fantastic opportunities to study or observe a spectacular trace fossil formed in unique conditions some 470 million years ago. Several geosites offer the possibility to literally walk upon an Ordovician seafloor (Figs. 5D–E). Within the context of the evolution of Earth’s Biosphere these are some of the earliest examples where the activity of animals thoroughly reworked the seafloor. In the framework of a paleoichnological research project to elucidate the paleobiological importance of Daedalus, the 800 km-long Armorican Quartzite Tourist Route will promote existing tourism related to the Appalachian-like quartzite landforms and their fossil sites and is already inspiring new sustainable businesses(Figs. 5F–H). This cross-border project, supported by scientific research on the complex architecture of Daedalus, is strengthening geotourism as an innovative and differentiating product for the whole region.

Figure 5. The Armorican Quartzite Route uniting by road Villuercas-Ibores-Jara and Naturtejo UNESCO Global Geoparks. A) The symbol of the route is inspired by the Daedalus spiral preservation on bedding planes. B) Orvalho GeoTrail at Naturtejo is one of the attractions that the Armorican Quartzite Route will connect. C) Road signs planned for the identification of the route. D) International Appalachian Trail panel with the Daedalus as identity symbol, Naturtejo. E) Via Ferrata “Walking on the Ordovician Ocean” at the Muradal mountain, Naturtejo. F and G) The Geocakes company, a partner of Naturtejo, with shop and cookies inspired by Daedalus. H) Training sessions with geoscientists in geosites for partner companies of Villuercas-Ibores-Jara Geopark.
Acknowledgments
The paper is part of the Cross-Border Interreg project REDTI, CBC PROGRAMME SPAIN-PORTUGAL 2007-2013 (POCTEP), which was extended to June 30th 2021. The fieldwork of Andrea Baucon was supported by the PALEOGIANTS project (University of Genova). The authors greatly appreciate the work of the reviewers Eamon Doyle and Alan Briggs.
Conflict of Interest
The authors have no known conflict of interest.
References
Barrera JM & Gil Montes J (2013). Guía de Geositios del Geoparque Villuercas-Ibores-Jara. Cáceres: Diputacón Provincial.
Barrera JM & Neto de Carvalho C (2018). The bridge over the Armorican Quartzite. Global Geoparks Network Newsletter 2018. 1: 8–9.
Bayet-Goll A & Neto de Carvalho C (2020). Architectural evolution of a mixed-influenced deltaic succession: Lower-to-Middle Ordovician Armorican Quartzite in the southwest Central Iberian Zone, Penha Garcia Formation (Portugal). International Journal of Earth Sciences. 109: 2495–2526.
Beuf S, Biju-Duval B, Charpal O, Rognon P, Gariel O & Bennacef A (1971). Les grès du Paléozoïque inférieur au Sahara. Sédimentation et discontinuités, évolution structurale d’un craton. Paris: Technip.
Bock MJ & Miller DC (1995). Storm effects on particulate food resources on an intertidal sandflat. Journal of Experimental Marine Biology and Ecology. 187: 81–101.
Cortijo I, Caballero J.L, Barrera JM, Gil Montes J & Palacios T (2016). Guía Turistica del Geoparque Mundial de la UNESCO Villuercas-Ibores-Jara. Cáceres: APRODERVI.
Delgado JFN (1885). Terrenos paleozóicos de Portugal: Estudo sobre os Bilobites e outros fósseis das quartzites da base do Systema Silurico de Portugal. Lisbon: Mem. Sec. Trab. Geol. de Portugal.
Desai BG, Shukla R & Saklani RD (2010). Ichnology of the Early Cambrian Tal Group, Nighalidar Syncline, Lesser Himalaya, India. Ichnos. 17: 233–245.
Desjardins PR, Mángano MG, Buatois LA & Pratt BR (2010). Skolithos pipe rock and associated ichnofabrics from the southern Rocky Mountains, Canada: colonization trends and environmental controls in an early Cambrian sand-sheet complex. Lethaia. 43: 507–528.
Droser ML (1991). Ichnofabric of the Paleozoic Skolithos Ichnofacies and the nature and distribution of Skolithos piperock. Palaios, 6: 316–325.
Durand J (1985). Le Grés Armoricain. Sédimentologie, Traces Fossiles, Milieux de dépôt. Rennes: Memoires et Documents du Centre Armoricain d’Etude Structurale des Socles. 3.
Gibert J, Ramos E & Marzo M (2011). Trace fossils and depositional environments in the Hawaz Formation, Middle Ordovician, western Libya. Journal of African Earth Sciences. 60: 28–37.
Gutérrez-Marco JC, Lorenzo S & Sá AA (2017). Fontanarejo (Ciudad Real): una localidad icnologica excepcional del Ordovícico Inferior en los Montes de Toledo meridionales. Geogaceta. 52: 47–50.
Gutiérrez-Marco JC, Rábano I, Sá AA, San José M, Pieren Pidal AP, Sarmiento GN, Piçarra JM, Durán JJ, Baeza E & Lorenzo S (2007). Public dissemination of knowledge regarding Ordovician geological and palaeontological heritage in protected natural areas of Iberia. Acta Palaeontologica Sinica. 46: 163–169.
Heward AP, Miller CG & Booth GA (2019). The Early Ordovician Middle Shale Member (Am3) of the Amdeh Formation and further evidence of conodont faunas from the Sultanate of Oman. Geological Magazine. 156: 1357–1374.
Lessertisseur J (1971). L’énigme du Daedalus (Daedalus Rouault, 1850). Ichnofossilia. Bulletin du Muséum National d’Histoire Naturelle, Sciences de la Terre. 20: 38–65.
Liang F, Liu JB & Zhan ZB (2012). Temporal distribution of piperocks in Cambrian and Ordovician: A coevolutionary process with changes of paleoenvironment. Science China, Earth Sciences. 55: 26–38.
López Caballero J, Barrera Martín-Merás JM & Cortijo Sanchez I (coord.) (2018). The Villuercas-Ibores-Jara UNESCO Global Geopark. Cáceres: Diputación de Cáceres.
Mángano MG & Buatois LA (2011). Timing of infaunalization in shallow marine early Paleozoic communities in Gondwanan settings: discriminating evolutionary and paleogeographic controls. Palaeontologia Electronica. 14(2): 9A, 21 pp.
Mángano MG & Buatois LA (2015). The trace-fossil record of tidal flats through the Phanerozoic: Evolutionary innovations and faunal turnover. Geological Association of Canada, Miscellaneous Publication. 9: 157–177.
McIlroy D & Garton M (2010). Realistic interpretation of ichnofabrics and palaeoecology of the pipe-rock biotope. Lethaia. 43: 420–426.
Neto de Carvalho C (2003). Fractal implications in spatial and evolutionary paleoecology of the Daedalus producer (ichnofabric analysis). [In Abstracts Book of] XIV International Congress on Mathematical Physics. Lisbon: World Scientific.
Neto de Carvalho C & Baucon A (2013) Multifractals and Capacity Dimension as measures of disturbance patch dynamics in Daedalus ichnofabrics. In Mathematics of Planet Earth (pp. 747–751). Berlin: Springer Lecture Notes in Earth System Sciences.
Neto de Carvalho C, Baucon A & Gonçalves D (2016). Daedalus mega-ichnosite from the Muradal Mountain (Naturtejo Global Geopark, central Portugal): between the Agronomic Revolution and the Ordovician Radiation. Comunicações Geológicas. 103: 59–70.
Neto de Carvalho C, Jensen, S, Palácios T, Barrera, J.M, Cortijo I & Caballero JL (2018). Daedalus: symbol of the bridge over the Armorican Quartzite uniting Villuercas-Ibores-Jara (Spain) and Naturtejo (Portugal) UNESCO Global Geoparks. [In Abstracts Book of] 8th International Conference on UNESCO Global Geoparks, “Geoparks and Sustainable Development” (pp. 196). Italy: Adamello Brenta Geopark.
Neto de Carvalho C & Rodrigues NPC (2002). Los Zoophycos del Bajociense-Bathoniense de la Praia da Mareta (Algarve, Portugal): Arquitectura y finalidades en régimen de dominancia ecológica. [In Abstracts Book of] Congresso Ibérico de Paleontologia (pp. 98–99). Spain: Univ. Salamanca.
Neto de Carvalho C & Rodrigues NPC (2003). Los Zoophycos del Bajociense-Bathoniense de la Praia da Mareta (Algarve, Portugal): Arquitectura y finalidades en régimen de dominancia ecológica. Revista Española de Paleontologia. 18: 229–241.
Noffke N (2012) Daedalus halli: a fossil biofilm harvester from the Lower Arenigian, Montagne Noire, France. [In Abstracts Book of] GSA Abstracts with Programs. 44(7): 500.
Poiré DG, Spalletti LA & del Valle A (2003). The Cambrian-Ordovician siliciclastic platform of the Balcarce Fm. (Tandilia System, Argentina): facies, trace fossils, palaeoenvironments and sequence stratigraphy. Geologica Acta. 1: 41–60.
Reineck HE (1963) Sedimentgefüge im Bereich der südlichen Nordsee. Abhandlungen der Senckenbergischen Naturforschenden. Gesellshaft. 505: 1-138.
Riedl RJ, Huang N & Machen R (1972). The subtidal pump: a mechanism of interstitial water exchange by water action. Marine Biology. 13: 210–221.
Rouault M (1850). Note préliminaire sur une nouvelle formation découverte dans le terrain silurien inférieure de la Bretagne. Bulletin Société Géologique du France. 7: 724–744.
Sá AA, Meireles C & Coke C (2002). Concentração maciça de Daedalus labechei (Rouault) (icnofóssil ordovícico) no Alto de Martim Preto (Guadramil-Bragança): património paleontológico a preservar e divulgar. [In Abstracts Book of] Congresso Ibérico de Paleontologia (pp. 138–139). Spain: Univ. Salamanca.
Sarle CJ (1906). Arthrophycus and Daedalus of burrow origin. Proceedings of the Rochester Academy of Science. 4: 203–210.
Seilacher A (1964). Sedimentological classification and nomenclature of trace fossils. Sedimentology. 3: 253–256.
Seilacher A (2000). Ordovician and Silurian Arthrophycid ichnostratigraphy. In Geological Exploration in Murzuq Basin (pp. 237–258). Elsevier Science B. V.
Seilacher A (2007). Trace fossil analysis. Berlin Heidelberg: Springer.
Thayer CW (1983). Sediment-mediated biological disturbance and the evolution of marine benthos. In Biotic Interactions in Recent and Fossil Benthic Communities (pp. 480–625). Plenum Press. New York.