Introduction
Rock strata that exhibit extraordinary fossils with exceptional preservation are called Konservat-Lagerstätten (Seilacher et al. 1985). Many Lagerstätten are known worldwide, including the famous Burgess Shale deposits of British Columbia, the Cambrian Chengjiang Fossil Site in China, the Devonian Great Canadian Lagerstätten, the Cretaceous Jehol Biota of China, the Cretaceous Santana Formation in Brazil and the Eocene Messel Pit Fossil Site in Germany. Some of these sites have been designated as UNESCO World Heritage Sites while others are part of the UNESCO Global Geopark network.
Fossils from the laminated limestones of the Luberon present a rich record of an early Oligocene terrestrial community with exceptionally preserved fauna and flora, displaying the anatomy of hard and soft tissues in a wide variety of organisms. The fossiliferous limestones were deposited in a large lake, which occupied the region during the early Oligocene (Ducreux et al. 1985) providing exceptional fossilization conditions. These fossils offer unique information about the early stages of the evolution of modern lineages and help to understanding the living environment of the Oligocene, around 30 million years ago.
These delicate fossils are vulnerable and fragile, facing different kinds of threats, including natural deterioration and collecting pressures. Since 1987, the Lagerstätte has been protected by the Luberon Geologic National Nature Reserve (Luberon Geologic NNR). The Luberon Geologic NNR is managed by the Luberon Regional Nature Park recognized as a UNESCO Global Geopark (UGGp).
We report here on the approaches to protect and promote this remarkable geoheritage. Through the efforts of the Luberon UGGp, the laminated limestone fossil sites and their fossils are effectively managed and valorized by scientific research and public education.
Geological Setting
The Apt-Manosque-Forcalquier Basin is one of continental basins that originated during the Oligocene in southern France in response to extensional tectonics that affected the European platform, leading to the formation of the West-European rift system and the Liguro-Provençal basin (Hippolyte et al. 1993). The filling of the basin is characterized by shallow water continental sedimentation (Nury 1988).
The laminated limestones of the Apt-Manosque-Forcalquier Basin that produce the fossils belong to the lacustrine Campagne-Calavon Formation. Based on biostratigraphic data obtained from gastropods (Cavelier et al. 1984), charophytes (Feist 1977), ostracods (Apostolescu & Guernet 1992) and mammals (Helmer & Vianey-Liaud 1970; Ducreux et al. 1985), this formation is dated as Rupelian (biozone MP23–24).
The main laminated limestones fossiliferous localities of the Campagne-Calavon Formation, protected by the Luberon Geologic NNR, are shown in Fig. 1. The localities around Céreste (Bastide Barbély, Bastide du Bois, Grivet), Viens (La Bonnette, Cavalier, Les Chapelins, Saint-Jean, Triclavel) and Vachères (Pichovet) extensively excavated by amateurs and scientists are best known for insects, isolated feathers, sub-complete skeletons and plant remains.
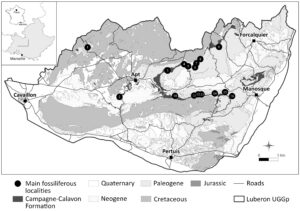
Figure 1. Geographical and geological setting of the early Oligocene Campagne-Calavon Formation and the main fossiliferous localities protected by the Luberon geologic National Nature Reserve. (1) Le Puy (Murs, Vaucluse); (2) Les Chapelins (Bonnieux, Vaucluse); (3) Grand Plan (Caseneuve, Vaucluse); (4) La Bonnette (Viens, Vaucluse); (5) Saint-Jean (Viens, Vaucluse); (6) Triclavel (Viens, Vaucluse); (7) Cavalier (Viens, Vaucluse); (8) Grand Banc (Oppedette, Alpes-de-Haute-Provence); (9) Pichovet (Vachères, Aubenas-les-Alpes, Revest-des-Brousses, Alpes-de-Haute-Provence); (10) Glorivette (Saint Martin de Castillon, Vaucluse); (11) Grivet (Céreste, Alpes-de-Haute-Provence); (12) Bastide Barbély (Céreste, Alpes-de-Haute-Provence); (13) Bastide du Bois (Céreste, Alpes-de-Haute-Provence); (14) Cayols (Montjustin, Alpes-de-Haute-Provence); (15) Le Rocher (Montfuron, Alpes-de-Haute-Provence); (16) La Violette (Montfuron, Alpes-de-Haute-Provence). SIT-PNR-PACA, M. Krebs.
The Campagne-Calavon Formation documents a quiet, calm, and shallow lacustrine environment characterized by periodic variations of salinity (Ducreux 1982; Lutz 1984). The fossil ichthyofauna consists of a mixture of freshwater and euryhaline taxa (Gaudant 1978a, b, 1997, 2013, 2015). The presence of articulated skeleton part of a ranid frog (UGGp collection, undescribed specimen) is evidence of freshwaters in the basin. Connections to the sea may have episodically occurred as indicated by the fossil jellyfishes (Nel et al. 1987a). The preservation of tiny morphological structures and soft parts suggest that microbial films played a role in fossilization, promoting rapid embedding of the organisms (Gall et al. 1990; Nel 1991; Nidergas et al. 2018; Iniesto et al. 2017).
An Oligocene Fossil-Lagerstätte
The fossil record of the Campagne-Calavon Formation has been known since the 19th century. The fossil fish that dominate the fauna were first mentioned by Sauvage (1880). The most abundant taxa include the perciform Dapalis macrurus (Fig. 2E) and the cyprinodont Prolebias goreti (Sauvage 1880; Gaudant 1978a, b, 1997, 2013, 2015).
Saporta (1891) studied the fossil flora and documented hundreds of species based on flowers, seeds and leaves remains. A diversity of plants ranging from ferns (Osmunda, Lygodium, Aspidium), gymnosperms and many angiosperms have been identified (Saporta 1891; Coquelle 1957; Thiébaut 1998).
Fossil amphibians are represented by an articulated skeletal remain of a ranid frog (UGGp collection, undescribed specimen). Fossil reptiles include an undescribed complete shell and other skeletal elements of a turtle (Luberon UGGp Geology Museum collection) an almost complete skull and postcranium skeleton of a crocodile (Diplocynodon ratelii) (Brinkmann & Rauhe 1998).
The Luberon is well-known for fossil birds, being among the few sites that yielded well-preserved partial skeletons as well as impressions of isolated feathers of Oligocene birds in Europe (Stemvers-van Bemmel 1984; Mourer-Chauviré 1996; Roux 2002; Mayr 1999, 2001, 2005; Louchart et al. 2008, 2011; Duhamel et al. 2020; Riamon et al. 2020). The bird record includes one of the most complete specimens of trogons (Mayr 1999, 2001), an exceptionally preserved almost complete specimen of a modern-type hummingbird (Louchart et al. 2008) and one of the earliest fossil passerine birds (Fig. 2A; Riamon et al. 2020).
Mammalian remains are rarer, with a few specimens known. These include a subcomplete skeleton of the artiodactyl Bachitherium cf. insigne (Fig. 2B; Geraads et al. 1987), as well as chiropteran material (Molossidae) and a rodent. The chiropteran remains consist of two nearly complete specimens currently under study, and an isolated forelimb referred to the molossid Tadarida described by Sigé (1971). The complete skeleton of a theridomyid rodent has been attributed to Pseudoltinomys gallairdi (Schmidt-Kittler & Storch 1985).
The entomofauna is rich and exceptionally well-preserved. Among the fossil insects, terrestrial groups are very dominant, with flies (Bibionidae) and weevils (Curculionoidea) being the most common groups; aquatic insects are rare (Oustalet 1874; Fabre 1914; Théobald 1937; Nel 1984, 1985, 1986, 1987a,b,c, 1991; Fernández-Rubio & Nel 2000; Nidergas et al. 2018; Claisse et al. 2019). The most represented taxa include dipteran bibionids, coleopteran curculionids and ground beetles (Carabidae) and grasshoppers (orthopteran acridids). Well-preserved Discalioides fossils have been interpreted as the remains of the discoid rigid pneumatophore-like structure of small jellyfishes (Nel et al. 1987).

Figure 2. Fossils from the early Oligocene laminated limestone deposits of the Luberon UGGp, southeastern France. A) Fossil passerine bird (Tyrannida, Passeriformes) from Revest-des-Brousses (Alpes-de-Haute-Provence), NT-LBR-014 (collection Nicolas Tourment, Marseille, after Riamon et al. 2020). B) Fossil ruminant Bachitherium cf. insigne (Mammalia, Artiodactyla) from Pichovet (Vachères, Alpes-de-Haute-Provence), PNRL-1992 (Luberon UGGp Geology collection). C) Fossil lepidoptera Lethe (?) corbieri (Nimphalidae) from Bastide du Bois, Céreste (Alpes-de-Haute-Provence), PNRL-1575 (Luberon UGGp Geology collection). D) Fossil pine cone (Plantae, Pinophyta) from Pichovet, (Vachères, Alpes-de-Haute-Provence), PNRL-1266 (Luberon UGGp Geology collection). E) Fossil fish Dapalis macrurus (Perciformes) from Le Grand Banc, Oppedette (Alpes-de-Haute-Provence), PNRL-1264 (Luberon UGGp Geology collection).
These delicate fossils, rarely preserved elsewhere, yield valuable scientific data offering unique insights into past terrestrial and lacustrine ecosystems. They increase our knowledge of biodiversity during the Oligocene, as well as the general paleoenvironmental context and stratigraphic evolution of Luberon. There are other known Oligocene Lagerstätten. Some of these, the late Oligocene Enspel Lagerstätte, the latest Oligocene deposits of Aix-en-Provence, the late Oligocene Riversleigh site and the latest Oligocene amber in Chiapas, have a high diversity of well-preserved fossils that add to our understanding of Oligocene life, but most of them document the latest part of the Oligocene. The Luberon laminated limestone deposits are unique in the diversity and significance of early Oligocene animals and quality of preservation. They are indicative of a warm subtropical climate (Châteauneuf & Nury 1995). They document the flora and fauna of southern France after the ‘Grande Coupure’, which represents a major terrestrial faunal turnover recorded in Eurasia associated with the overall climate shift at the Eocene–Oligocene transition.
Protection and Geoconservation
Protection of geological heritage as part of the natural heritage dates back to the beginning of the twentieth century in France. Nature reserves were created in 1976 as a legal tool to protect the geological heritage (Avoine 2012; De Wever et al. 2014).
Since 1987, the Luberon Geologic NNR has protected 28 paleontological sites. The reserve area was extended in 1996 to include, along with other fossiliferous outcrops, most of the Campagne-Calavon laminated limestones. The nature reserve regulations formulate administrative, scientific and organizational conditions for excavations. Extraction and collection of fossils and minerals are prohibited without prior special authorization.
Until the reserve was created, uncontrolled fossil collecting and trade was taking place. Some of the sites have been particularly exploited by fossil traders.
In 1981, the almost entire skeleton of a small herbivorous mammal was discovered by a private collector at the Pichovet (Vachères, Alpes-de-Haute-Provence) locality (Fig. 2B). This represents the only known whole specimen of the primitive ruminant Bachitherium (Geraads et al. 1987). The fossil was sold to a German collector in 1984 and remained in his possession before he decided to sell it in 1991. The Luberon Regional Nature Park (management structure of the Luberon Geologic NNR and the Luberon UGGp) organized funding and bought the specimen to ensure that it would be legitimately available for study and public display. This fossil now belongs to the Luberon UGGp Geology Museum and is on display at the regional Pierre Martel Museum of Vachères.
The UGGp Geology Museum, located in the city of Apt (Vaucluse), consists of two exhibition rooms and contains scientifically important collections of more than 4000 fossil animal and plant specimens. Some of the earlier fossil collections from the laminated limestones of Luberon are found in national and international museums in Paris (Museum National d’Histoire Naturelle), Frankfurt (Senckenberg), Lyon (Université Claude Bernard), Aix-en-Provence (Muséum d'Histoire Naturelle) while many other spectacular and important specimens are held in private collections.TheUGGp Geology Museum is dedicated to educating the public through the presentation of the Luberon territory geological history and the display of its paleontology collection. It provides an adequate storage place for the fossils, ensures their proper curation and documentation as well as their accessibility and long-term security.
As a geopark, the Luberon UGGp promotes the protection, management and valorization of its world-class paleontological heritage. Today, there are still important undescribed fossils in private and public collections. One of the priorities is to inventory and improve access to these important fossils.
Compared to other famous Paleogene Lagerstätten like the Messel Fossil Pit in Germany or the Eocene Green River Formation in the United States, the Luberon Lagerstätten is little known. Ongoing field research, led by the Museum National d’Histoire Naturelle (Paris), focuses exclusively on fossil insects. Developing scientific research, field training and outreach are keys to better understand and promote this paleontological heritage. The long-term conservation and valorization strategy requires us to strengthen academic research and field projects. Encouraging educational activities, notably by using these fossils to explore ancient climate changes, public outreach including paleontology and geology talks, participation in fossil quarry collections is also essential to share and preserve this rich heritage.
Conflict of Interest
The authors declare that they have no competing interest
Acknowledgments
The authors express sincere thanks to Christine Balme, the former curator of the Luberon Geologic NNR, who spent most of her career protecting and promoting the geoheritage of Luberon.