Results and discussion
One of microalgae potential’s benchmarks as biomass fuel feedstock is determined by the abundance of the biomass so the availability will be assured. Therefore, the biomass productivity of algae in terms of growth rate is one of the most significant parameters that must be examined.
The biomass yields of
N. oculata
during 7-day cultivation period were depicted in Fig.
1
. These results showed that
N. oculata
multiplied its biomass rapidly, starting at around 2 × 10
6
cells/ml on day 1 to approximately 12 × 10
6
cells/ml on day 7. It meant the average increase in biomass was approximately 1.5 × 10
6
cells/ml/day, higher than that of
Selenastrum
sp., which increase in cell number per day was about 1.6 × 10
4
up to 2.2 × 10
4
cells/ml/day [
41
]. If the productivity of
N. oculata
was expressed in growth kinetics, then it had specific growth rate of 0.27/day and doubling time of 2.59 days as shown in Table
2
. Its growth kinetics revealed that
N. oculata
biomass productivity was higher than
Dunaliella salina
[
42
],
S. obliquus
[
43
], and
C. vulgaris
[
44
], which their growth rates were 0.18, 0.22, and 0.14/day, and their doubling time were 3.85, 3.15, and 4.95 days, respectively. This result was important to note by considering the
N. oculata
had been cultivated in the traditionally natural open pond as illustrated in Fig.
2
a. The products of biomass sediment, the dried biomass chunk and the biomass powder of
N. oculata
, were shown at Fig.
2
b–d, respectively. By fitting the curve of Fig.
1
, the growth pattern of
N. oculata
could be estimated as an order 3 of a polynomial equation as
Y
= 2,018,570 − 736,666.67
X
+ 783,095.24
X
2
− 66,666.67
X
3
with the fitting degree (
R
2
) of 0.999, where
Y
and
X
were cell density and cultivation time, respectively.
The cell density of
Nannochloropsis oculata
during 7-day cultivation period. Relative standard error of means were below 5 % for all situations The growth kinetics parameters and properties of
Nannochloropsis oculata Parameters/properties Values Specific growth rate (/day) 0.27 Doubling time (day) 2.59 Lipid content (%) 11.44 EDX analysis (wt%) C 28.32 O 43.80 Na 2.04 Mg 13.16 Al 0.92 Si 1.60 Cl 1.97 Ca 8.20 Proximate analysis (wt%) 3.99 VM (db) 67.45 FC (db) 8.08 24.47 HHV analysis (MJ/kg) 16.80 The cultivation pond (
a
), the biomass sediment (
b
), the dried biomass chunk (
c
), and biomass powder (
d
) of
Nannochloropsis oculataFig. 1
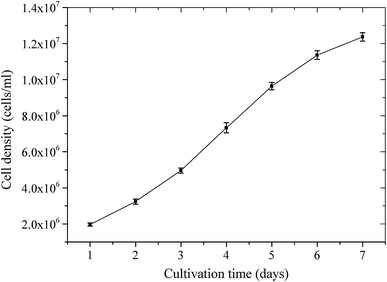
Table 2
Fig. 2

The properties of algal biomass as a biofuel source were taken into consideration in this study. These properties varied widely, depended on the kind of algae, environmental culturing, cultivation periods, and harvesting conditions. Hence, investigating these properties was essential to determine the proper combustion technology.
Figure
3
showed the EDX spectrograms of
N. oculata
biomass for determination of its compositions as compiled in Table
2
. Generally, the investigated
N. oculata
biomass has low carbon, high oxygen, and high inorganic elements compared to the result of Patil et al. [
45
]. These compositions had an impact on biomass combustion and the residual characteristics. They were also needed for estimating the air required for the combustion process and for predicting the released gases. Carbon was oxidized during combustion by exothermic reactions and formed CO
2
. The organically bound of oxygen is released as the result of thermal processes, and it supplies a part of the overall oxygen needed for the combustion reactions [
14
].
EDX spectrograms of
Nannochloropsis oculata
biomassFig. 3
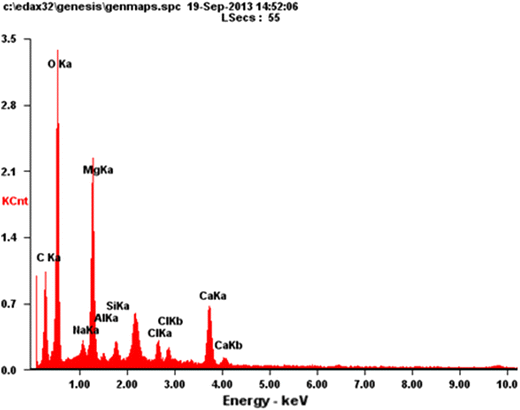
The high amount presence of inorganic materials in this biomass, especially magnesium, calcium, sodium, silicon, and chlorine needs to be considered because they frequently results in slagging and fouling problems in furnaces. The reaction of alkali metals with silica presented in the ash can lead to the formation of low-melting silicates in fly ash particles yielding a sticky, mobile liquid phase, which causes blockages of airways in the furnace and boiler plant [ 46 ]. Cl vaporized during combustion, forming HCl, Cl 2 , and alkali chlorides. Alkali and alkaline earth chlorides are condensed in the boiler section as fly- ash particles or on the heat exchanger surfaces when flue gas temperature decreases [ 47 ]. Parts of the Cl were bounded in the fly ash while the rests were emitted as HCl in the flue gas. HCl emissions could play in the corrosion of metal component plant such as boilers and air pollution control devices.
The TG curve for determining the proximate value was shown in Fig.
4
. The values of moisture (
M
), volatile matter (VM), fixed carbon (FC), and ash (
A
) were directly determined from the TG curve by the difference while higher heating value (HHV) of the biomass was estimated by Eq. (
3
). All of these results were tabulated in Table
2
.
TG curve for determining the proximate value.
a
Mass loss (%) vs. time (min) and
b
temperature setting (°C) vs. time (min)Fig. 4
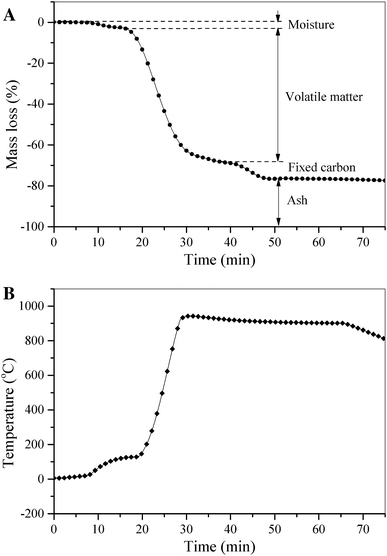
As shown in Fig. 4 , the first mass loss, which was indicated by the reaching of the first constant mass line, corresponded to moisture loss. The N. oculata moisture content was 3.99 %, which seemed to be a potential candidate for direct combustion. The moisture content influenced the combustion behavior and the amount of energy to evaporate water. The water content in the fuel had to be released before the first step combustion starting; hence high moisture content of biomass meant more energy for evaporation, and subsequently for heating the water vapor. Removal of water in the biomass would decrease the maximum possible combustion temperature and overall system efficiency. Aside from moisture evaporation that consumed a great deal of heat, it also caused the evaporation time to be extended during the combustion process. Hence, it resulted in the decay of ignition and affected the necessary residence time of biomass fuel in the combustion chamber before gasification and combustion taking place. Consequently, the high moisture fuel contents would require the larger combustion chambers and would result in an expansive boiler. Moreover, the amount of moisture also affected the burning reaction rate; thus, it influenced the generated gas emission as NO and CO, as well. In the devolatilization process, due to evaporated moisture moves away from the devolatilization front, the amount of volatile determined the reaction zone thickness [ 48 ], and it influenced the heat transfer to the devolatilization zone which affected the devolatilization rate. Thus, the moisture content was very important fuel parameter, and investigating this parameter was required to adjust the temperature control system of the furnace properly.
In line with the rising temperature, a major part of biomass organic component experienced thermal cracking then decomposed and volatilized to be the volatile matter. In inert gas, the volatile is released at temperature up to 900 °C [ 39 ]. For microalgal biomass, it was especially resulted from the thermal cleavage of proteins, carbohydrates, and lipids. Table 2 showed that N. oculata volatile matter content was 67.45 %, lower than Chlorella sp. MP-1 that was 77.45 % [ 49 ] and higher than C. vulgaris that was 55.37 % [ 50 ]. The amount of biomass volatile matter strongly influenced the thermal decomposition and the combustion behavior. As it had been previously stated that the biomass, which had a high amount of volatiles, would be degraded as a result of the heating process. A significant part of the biomass was vaporized earlier before homogeneous gas phase combustion reactions took place and subsequently the remaining char undergone heterogeneous combustion reactions. Because of volatile matter was the reactive substance, hence, more volatile meant easier biomass to be burned. The aforementioned phenomenon would determine both of the ignition and burnout time of volatiles in the first stage of combustion processes, as well as the char combustion in the second stage. Moreover, in the context of co-combustion with other solid fuel, higher volatile could create larger fuel-rich regions in the near-burner region. The fuel-rich regions were very critical area. They were used for flame stabilization but, the formation of undesired prompt NO x , H 2 S, and soot could occur in this zone, as well [ 51 – 53 ]. Hence, finding out its amount is necessary to adjust the sufficiency of air supplied in the zone where volatile is released to improve combustion efficiency. Furthermore, the appropriate composition of supplied air in the released volatile zone will affect the formation of soot and NO emissions as discussed earlier by Liu et al. [ 54 , 55 ].
The fixed carbon content, which was the mass left over after the releases of volatiles excluding the ash and moisture contents, produced a char and was burned as a solid material in the combustion system. The N. oculata fixed carbon content was 8.08 %. This content was lower than both of C. vulgaris [ 50 ] and Chlorella sp. MP-1 [ 49 ], which were 34.35 and 16.95 %, respectively. The volatile matter and fixed carbon contents significance were that they provided a measure of the ease which the biomass could be gasified and oxidized in the combustion process. The N. oculata biomass, which had high volatile matter and low fixed carbon contents, was highly reactive fuel and would give a faster combustion rate during the devolatilization phase. This amount of fixed carbon was also needed to estimate the heating value of biomass, and it was acted as the main heat generator during the burning process.
The final residual material after the combustion process of biomass was ash, and it formed a standard measurement parameter for solid fuels. Table 2 indicated that N. oculata ash content was 24.47 %, which was very high, four times higher than Chlorella sp. MP-1 that was 6.36 % [ 49 ]. It was in accordance with the EDX analysis results, which showed the inorganic components content of N. oculata was high, as well. Inorganic components in biomass had a direct influence in the ash formation. Ash-forming elements exist in biomass as salts [ 14 ], bounded in the carbon structure, or they present as mineral particles from cultivation environment and introduce into the biomass fuel during harvest. In the combustion process, the part of the ash-forming compounds in biomass was vaporized and released to the gas phase while non-vaporized ash compounds that were left over in the char might melt and coalesce inside and on the surface of the char. The vaporized compounds would either be condensed or reacted on the surface of pre-existing ash particles in the flue gas when its temperature decreased. All of these could contribute to bed agglomeration, heat transfer surface fouling, and system corrosion. Furthermore, in the combustion process, the ash resulted from char burning would be formed as the layer surrounding their surface; hence, it inhibited the oxygen diffusion during char combustion. It affects both burning rate and mass loss rate, as well [ 56 ]. To address the problem of N. oculata ’s high ash content during thermal conversion processes, it might suitably blend this biomass in terms of co-combustion with other feedstocks to obtain the overall low ash content. One of the low ash biomass was rice straw, which its ash content was around 0.1–0.7 % by dry weight [ 57 ]. It might appropriate be blended with N. oculata to be mixed biomass fuel feedstock. However, their characteristics have not been studied yet.
Higher heating value was a total of heat yielded by the complete combustion of a unit quantity of fuels, including the latent heat contained in the water vapor. Therefore, it represented the maximum amount of energy which was potentially recoverable from a given biomass source. Based on the proximate result, HHV of N. oculata was 16.80 MJ/kg. This value was lower than both of C. vulgaris and C. minutissima that were 18 and 21 MJ/kg, respectively [ 9 ]. According to Illman et al. [ 9 ] the heating value of algae is in correlation with the lipid content of their biomass rather than with other components such as carbohydrates and proteins. As a comparison, the lipid content of N. oculata in this work was about 11 %, whereas C. vulgaris and C. minutissima lipid content were 18 and 31 %, respectively [ 9 ].
The FTIR spectrum of
N. oculata
biomass and its residue were shown in Fig.
5
. For the biomass, it could be observed fifteen distinct transmission bands over the wave number range of 4,000–400/cm. These bands were assigned to specific molecular groups and were tentatively identified on the basis of biochemical reference standards and published FTIR spectra as quoted in Table
3
. The cellular macromolecules (proteins, lipids, and carbohydrates) could be identified by their distinct transmittance in different frequency regions. The “carbohydrate band spectra” was characterized by weak and medium features at around 1,101–839/cm due to C–O and C–O–C stretching, Si–O stretching of siloxane silicate frustules, Si–OH bond stretching and bending of silanol. The “protein band spectra” was presented as less pronounced band at about 1,242/cm owing to asymmetric stretching of phosphodiester P=O. The spectrum of 1,435/cm related to CH
2
and CH
3
bending of methyl and C–O stretching of carboxylic group. The band at around 1,504/cm was amide II band spectra mainly
(C–H) and
(N–H). The medium amide I band at around 1,811–1,599/cm was primarily for C=O stretching of esters. The less pronounced band at 2,523/cm and around 3,659–3,003/cm because of amine salts –NH
3
+
stretching and N–H stretching of amide A, respectively. The bands associated with “lipid band spectra” were indicated by two weak peaks at around 2,940–2,835/cm related to CH
2
stretching of methylene and a strong peak at 1,788/cm as a result of C=O stretching of esters. The aforementioned bands spectra have specified the presence of the principal components in the
N. oculata
, namely carbohydrate, protein, and lipid. These components derived from various microalgae cell structure, such as a cell wall, plasma membrane, chloroplasts, mitochondrion, etc. Each cell structure substance would be progressively destroyed during the thermal processes when the temperature reached its decomposition temperature, and part of it released and formed volatile matter, whereas the remaining substance formed char as a solid material. Both of volatile and char would be burned through the combustion processes. The last rests after their complete burning processes were ash as residual materials.
FTIR spectra of
Nannochloropsis oculata
biomass and its residue Tentative assignments of bands found in the FTIR spectra of
Nannochloropsis oculata
biomass Band Main peak (/cm) Wave number range (/cm) Typical band assignment References 1 3,694 3,759–3,676 Water [ 2 3,645 3,659–3,636 Water [ Protein 3 3,343 3,512–3,003 Water [ Protein 4 2,916 2,940–2,897 Lipid [ 5 2,849 2,860–2,835 Lipid [ 6 2,523 2,571–2,509 Protein [ Superimpossed (O–H) and –NH3+ stretching 7 1,788 1,811–1,765 Lipid (fatty acids) [ Protein 8 1,651 1,692–1,599 Protein [ Amide I band mainly 9 1,504 1,545–1,460 Protein [ Amide II band mainly DNA (cytosine) In-plane double bond base vibrations, which include C=C, C=N and C=O stretching 10 1,435 1,445–1,416 DNA (Adenine) [ Protein Carboxylic group 11 1,242 1,312–1,204 DNA-phospholipids Phosphodiester of nucleic acids and phospholipid [ Antisymmetric 12 1,082 1,101–1,070 Phospholipids, DNA and RNA [ Carbohydrates Siloxane, silicate frustules 13 1,001 1,020–957 Siloxanes [ Silanes Si–OH bond stretching Carbohydrates RNA–DNA Ribose 14 880 893–870 Polysaccharides of the cell wall [ β- Silanol 15 854 866–839 Polysaccharides of the cell wall [ α- SilanolFig. 5
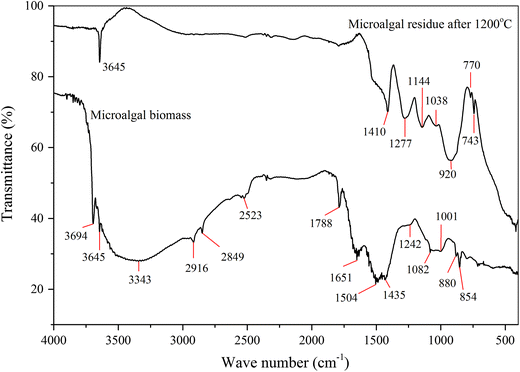
Table 3
Microalgal residue bands spectra were characterized by a strong feature at 3,645/cm related to O–H stretching of adsorbed water by silica presented in the ash, and strong, medium, and weak features at the fingerprint area associated with the rest of the inorganic component which was left in the ash. The band spectra between 1,490 and 1,410/cm correlated with calcium carbonate, mainly due to antisymmetric CO 3 – stretching [ 58 ]. The features at around 1,200–900/cm region associated with the silica structure, especially due to Si–O stretching modes [ 59 ]. The peaks below 800/cm were vibrations of Ca–O and Mg–O [ 60 ]. The strong band at around 400–500/cm is caused by Si–O or Al–O bending vibrations [ 61 ]. The elements of ash content were critical parameter in that case of byproduct handling and utilization, as well as they have an impact on the overall processing cost of biomass conversion. However, the ash characterization, other than FTIR that have been previously stated, was not performed in this research, and it will be extensively investigated in the future studies.
Figure 5 revealed the progressive degradation intensity in the spectra of biomass residue after 1,200 °C, mainly at around 3,590–1,630/cm. The declined intensity was caused by the loss of protein and lipid components of biomass during thermal processes. The feature changes at fingerprint area associated with protein degradation of biomass mainly amide II. It also correlated with degradation of cell wall structures including their fibrillar component that composed of polymer of β- d -glucose and their amorphous mucilaginous material which constituted polysaccharides, lipids, and proteins.